Le métabolisme énergétique (2)
1 La régulation des apports
Nous savons que le métabolisme des cellules de l'organisme nécessite un apport de nutriments régulier, par exemple pour le renouvellement cellulaire. D'autre part, des besoins énergétiques irréguliers existent au niveau de l'organisme : production de chaleur lorsqu'on s'éloigne de la température de neutralité thermique, activité musculaire, etc. Cette énergie, nous l'avons vu précédemment, dérive aussi des molécules organiques issues des aliments, et ce par un mécanisme de conversion de leur énergie potentielle en ATP. Cependant, de même que la quantité de nutriments utilisée est variable, leurs apports aussi (Ils sont par exemple intenses lors des repas.) Il faut donc réguler ces apports pour parvenir à un équilibre.
La nécessité de la régulation ne se trouve pas que dans ces considérations d'ajustement quantitatif. En effet, un facteur qualitatif intervient. Certaines cellules ont une exigence absolue et permanente en glucose (elles sont dites glucodépendantes): citons les hématies ou les neurones. Quand bien même l'apport global de nutriment resterait constant au cours du temps, il faudrait en outre que la proportion de glucose reste fixe. Ceci est manifestement irréalisable. Une simple observation nous confirme l'existence du phénomène de régulation des apports en glucose. Alors que les ingestions d'aliments sont irrégulières, le taux de glucose dans le sang (appelé aussi glycémie) reste quasiment constant : ~1g /L le matin à jeûn, ~1, 2 g / L après un repas riche en glucides.
1.1 Mécanisme de régulation des apports en glucose
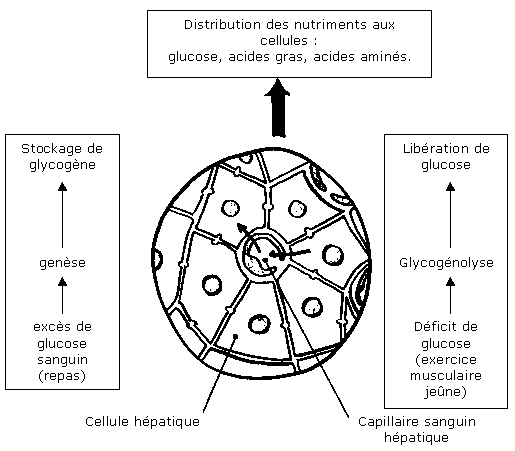
L'organe essentiel est ici le foie. Sa structure richement capillarisée permet l'existence d'une grande surface d'échanges entre cellules hépatiques et système sanguin. Notons que le sang provenant des anses intestinales, très riche en nutriments, traverse le foie. On peut dégager deux mécanismes hépatiques principaux relatifs à l'ajustement de la glycémie :
- Le foie est l'organe de stockage des excès de glucose. Pour cela, il forme une macromolécule glucidique: le glycogène grâce à un processus nommé glycogénogenèse. Le glucose peut aussi être insuffisant dans le milieu intérieur. Il existe donc une réaction inverse à la glycogénogenèse qui permet le retour du glucose dans le sang. Elle se passe en deux temps. En premier lieu, une glycogénolyse donne du glucose-phosphate à partir du glycogène. Or, celui-ci ne peut pas passer directement dans le sang (son groupement phosphate l'empêche de traverser la membrane des cellules). Il doit subir, dans une deuxième étape, une déphosphorylation placée sous l'action catalytique d'une enzyme hépatique. Par l'action conjointe de ces deux processus métaboliques, la concentration sanguine du glucose est efficacement régulée.
- Le foie peut également libérer du glucose qui ne provient pas des réserves en glycogène: c'est la néoglucogenèse. Celle-ci s'effectue, en présence d'enzymes spécifiques, à partir de précurseurs non glucidiques tels que certains acides aminés, les triglycérides ou encore l'acide lactique.
1.2 Activité musculaire et régulation
Les cellules musculaires ont principalement besoin pour leur métabolisme de glucose-phosphate et d'acides gras. D'autre part, le tissu musculaire est capable de stockage :
- Des réserves glucidiques sont constituées sous forme de glycogène.
- Les triglycérides constituent des réserves lipidiques. Leur synthèse se fait à partir de glycérol et d'acides gras. Bien sûr, ils redonnent ces composés une fois hydrolysés.
Lors de l'activité musculaire, divers phénomènes se produisent. L'absence d'enzymes permettant la déphosphorylation du glucose-phosphate dans la cellule musculaire empêche le passage de glucose dans le sang. Lorsque l'activité se prolonge, les réserves glycogéniques finissent par être épuisées. Cette déperdition est heureusement palliée par le recours aux acides gras qui proviennent soit de l'hydrolyse des triglycérides musculaires, soit de la circulation sanguine.
Notons enfin que, pendant son activité, la cellule musculaire peut égaalement oxyder du glucose. Celui-ci est alors directement importé du milieu sanguin et dérive des glycogénolyse et néoglucogenèse hépatiques. Cependant, cette voie est peu utilisée pour deux raisons. D'une part, le glucose traverse très lentement les membranes des cellules musculaires. D'autre part, une utilisation trop massive de glucose sanguin pourrait faire baisser la glycémie (hypoglycémie), ce qui est extrêmement dangereux pour le système nerveux (une forte hypoglycémie provoque un coma puis la mort).
2 Activité musculaire et processus énergétiques
2.1 La double nature des fibres musculaires
L'ATP est présent dans les cellules musculaires en très petites quantités. Il doit donc être restauré par l'intermédiaire de voies métaboliques particulières qui sont au nombre de trois. L'une de ces voies est aérobie (= elle utilise de l'O2): c'est la respiration. Or, chaque individu possède des capacités cardiaques et respiratoires propres, de même que ses fibres musculaires ont des caractéristiques précises. Ces facteurs font que la consommation en O2 par unité de temps est limitée: on appelle VO2 max. la valeur maximale qui peut être atteinte. Celle-ci correspond, pour une fréquence cardiaque maximale, à 10 à 12 fois la consommation de repos. Il faut donc, lorsque VO2 max. est atteint (c'est le cas pour des exercices courts et intenses), qu'existent d'autres voies métaboliques qui n'utilisent pas de dioxygène. On qualifie ces voies métaboliques d'anaérobies. Celles-ci sont de deux sortes: fermentation lactique et fermentation alactique.
L'existence de ces trois voies métaboliques a une implication anatomique. Les fibres musculaires ont en effet une double nature :
- Les fibres blanches fonctionnent surtout de manière anaérobie. Leur contraction est rapide mais elles sont vite fatigables. Ce sont elles qui sont utilisées dans les exercices brefs
- Les fibres rouges, principalement aérobies, sont au contraire lentes mais moins fatigables. On les rencontre dans les muscles développant une activité prolongée.
2.2 Etude des processus chimiques
Etudions maintenant en détail les processus chimiques, libérateurs d'énergie, mis en jeu dans ces trois voies métaboliques.
- La voie anaérobie alactique, dans les fibres rapides et fatigables, utilise un composé
phosphorylé énergétique stocké dans les tissus musculaires: la phosphocréatine. La production
d'ATP se fait selon une réaction réversible:
phosphocréatine + ADP <-> créatine + ATP.
C'est donc l'hydrolyse de la phosphocréatine qui libère l'énergie permettant la phosphorylation de l'ADP en ATP. Les réserves en phosphocréatine étant faibles, cette voie métabolique est privilégiée dans les exercices de courte durée mais de forte puissance (ex. lancer du poids, sprint). - La fermentation lactique est une voie anaérobie. Cette voie métabolique intervient dans le cadre
d'exercices intenses, relativement courts (par exemple un 800 mètres) et prend place dans le cytosol
des fibres rapides. Les réactions en jeu font intervenir le glucose issu d'une glycogénolyse au niveau
des tissus musculaires.
La chaîne réactive est la suivante:
glucose-phosphate -> acide pyruvique + ATP, puis : acide pyruvique -> acide lactique - La respiration est mise en place pour les exercices de longue durée, au niveau des fibres
lentes, riches en mitochondries. En effet, l'acide pyruvique, issu de la dégradation du
glucose-phosphate, est complètement oxydé au niveau des mitochondries pour donner de l'eau
et du dioxyde de carbone. La réaction globale donne :
glucose (C6H12O6) + 6O2 -> 6 CO2 +6 H2O + 38 ATP.
Le glucose provient d'abord des réserves de la cellule ; du glycogène est décomposé, et des triglycérides sont transformés en glucose. Puis le glucose est prélevé dans le sang, depuis les réserves du foie en glycogène et en acides gras.
Au cours de l'effort, l'existence de ces voies métaboliques va expliquer la variation de la consommation de dioxygène. Au début de l'activité musculaire, l'O2 effectivement consommé est inférieur à celui nécessaire à la réalisation de l'exercice. Ce déficit s'explique par des ajustements physiologiques et biochimiques précédant l'établissement de la respiration cellulaire. Après l'exercice, la consommation effective d'O2 est au contraire plus élevée que celle habituelle de repos. Cet excès appelé EPOC (excès post-exercice d'oxygène consommé) est utilisé pour partie dans la reconstitution des stocks de phosphocréatine.
Mots-clefs :
- ATP, glucose, glycogène, muscle,
- phosphocréatine ; fermentation lactique ; respiration.