Le métabolisme énergétique (1)
1 Dépenses énergétiques et métabolisme
1.1 Les dépenses énergétiques
Nous avons vu dans les chapitres précédents que pour leur activité, par exemple la synthèse de nouvelles molécules organiques, tous les êtres vivants ont besoin d'énergie. Celle-ci provient de la dégradation de substrats organiques. Le phénomène de la respiration étant la principale source de production énergétique, son étude permet de comprendre les besoins énergétiques d'un individu. En effet, on peut facilement mesurer une intensité respiratoire (volume d'oxygène absorbé par unité de masse et par unité de temps). De là, si on connaît la quantité d'énergie correspondante à l'absorption d'un litre d'oxygène utilisé pour l'oxydation d'un substrat donné (ou coefficient thermique de l'oxygène), on calcule directement la dépense énergétique effective du sujet.
On montre alors que, bien qu'étant continu, le processus de dépense énergétique a une intensité est variable qui est soumise à plusieurs facteurs :
- A une échelle cellulaire, on observe tout d'abord que la dépense énergétique est différente selon le type de cellule étudié (on mesure pour cela les intensités respiratoires pour différents tissus). C'est par exemple le cas entre cellules chlorophylliennes et cellules animales.
- On constate que l'intensité respiratoire augmente avec l'activité musculaire. On peut donc relier dépense énergétique et activité musculaire.
- Chez les homéothermes (ou endothermes), nous savons que le corps doit être maintenu à une température interne constante. Il s'agit bien sûr d'un phénomène énergétique dont les modalités sont traduites par le phénomène de la respiration. Ainsi, l'intensité respiratoire augmente lorsque l'organisme s'éloigne d'une température dite de neutralité thermique, qui correspondant à la dépense d'énergie minimale. En d'autres termes, l'organisme dépense de l'énergie pour lutter contre le froid ou la chaleur.
1.2 le métabolisme de base
Si on considère que le métabolisme est l'ensemble des différentes dépenses énergétiques, on peut alors postuler que ce dernier varie selon les facteurs énoncés ci-dessus. Il est aisé de définir un métabolisme dit "de base" en fixant des conditions standard. Celles-ci sont la neutralité thermique, le repos musculaire (le sujet reste allongé) et le repos digestif (la dernière ingestion d'aliments date de 12 heures ou plus). On exprime les dépenses énergétiques irréductibles, notamment le mouvement respiratoire, le mouvement cardiaque et les opérations de renouvellement des tissus.
En d'autres termes, le métabolisme basal est la dépense énergétique la plus faible d'un individu en 24 heures. Elle correspond à une activité minimale et à l'entretien des structures de l'organisme.
Si nous rapportons maintenant cette grandeur à une unité de surface corporelle, et ce afin de tenir compte de la corpulence du sujet à laquelle sont intimement liés les échanges thermiques, on arrive au résultat suivant: le métabolisme de base est indépendant de l'âge du sujet, de son sexe et de son état de santé. Mieux, les valeurs inter-espèces sont très voisines.
2 Energie et ATP
Le métabolisme s'exprime, au niveau cellulaire, par un ensemble de réactions chimiques. On peut classer celles-ci en deux familles distinctes:
- Les réactions dites exergoniques (ou exoénergétiques) sont spontanées et libèrent de l'énergie ['-ergon-' : énergie, 'ex-' : vers l'extérieur].
- Les réactions dites endergoniques (ou endoénergétiques) nécessitent obligatoirement un apport d'énergie pour se dérouler ['endo-' : vers l'intérieur].
La présence de ces deux types de réactions dans la cellule permet un fait essentiel : les réactions exergoniques peuvent fournir l'énergie nécessaire à la réalisation des réactions endergoniques.
Les conditions suffisantes pour qu'un tel processus existe sont simples: la quantité d'énergie libérée doit être au moins égale à celle consommée et il doit exister entre les deux réactions un intermédiaire commun, qui capte l'énergie de la réaction exoénergétique et la redonne au moment voulu pour que la réaction endoénergétique puisse avoir lieu. Cet intermédiaire entre réactions exoénergétiques et endoénergétiques, c'est l'adénosine triphosphate ou ATP.
L'ATP est un ribonucléotide, composée d'une base azotée (l'adénine), d'un sucre (le ribose) et de trois groupements phosphate enchaînés. une enzyme spécifique détache un groupement phosphate terminal de la molécule d'ATP ; or cette liaison phosphate est très riche en énergie (30 kJ / mol), et cette énergie peut être exploitée pour une réaction endoénergétique.
La réaction est la suivante :
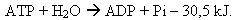
(un signe - devant une énergie signifie qu'elle est libérée lors de la réaction ; au contraire un
signe + indique que l'énergie doit être apportée pour que la réaction se fasse)
L'ADP (adénosine diphosphate) et le phosphate inorganique (Pi) peuvent redonner de l'ATP grâce à une enzyme spécifique (l'ATP-synthétase). Cette synthèse d'ATP est réalisée grâce aux dégradations des métabolites pendant la respiration ou pendant les fermentations ; en effet ces réactions libèrent l'énergie nécessaire à la reformation de l'ATP depuis l'ADP et le Pi.
Notons que l'énergie en excès est libérée sous forme de chaleur, qui ne peut être utilisée comme intermédiaire énergétique dans la cellule: ce sont des pertes.
Ainsi, la molécule d'ATP n'est pas une source d'énergie mais un intermédiaire énergétique entre les différentes réactions chimiques au sein de la cellule. Le fait qu'elle soit constamment hydrolysée et régénérée explique qu'elle est toujours présente dans le milieu cellulaire en très petites quantités.
3 Notion de voie métabolique
3.1 Fermentations et respiration
Les opérations de synthèse des molécules organiques (anabolisme) et de leur dégradation (catabolisme) sont réalisées par des chaînes complexes de réactions chimiques. On nomme celles-ci voies métaboliques.
L'ensemble des réactions d'une voie métabolique peut s'expliquer par un mécanisme d'oxydoréduction. L'oxydation est une réaction de perte d'électrons, la réduction est un gain d'électrons (attention !). Dans une réaction d'oxydoréduction, les électrons perdus pendant l'oydation sont utilisés pour la réduction : ces réactions s'effectuent toujours en couple.
On rencontre ainsi dans un premier temps des réactions d'oxydation catalysées par des enzymes spécifiques: les déshydrogénases. Les électrons ainsi libérés sont captés (réduction) par des molécules particulières, les transporteurs.
On peut décrire deux voies métaboliques principales :
- Les fermentations se déroulent dans le cytosol, et sont le plus souvent anaérobies : elles se
déroulent sans avoir besoin de dioxygène.
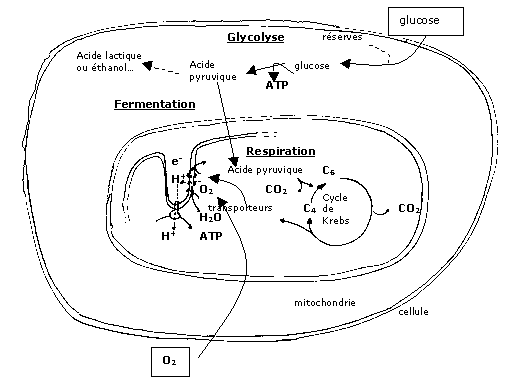
Elles comportent un petit nombre de réactions: ce sont des voies métaboliques courtes. Elles sont donc rapides mais la dégradation du substrat (par oxydation) reste incomplète. Leurs résidus organiques sont divers (par exemple : éthanol, ou acide lactique…) et intimement liés à l'équipement enzymatique de la cellule considérée. - La respiration, au contraire, est une voie métabolique longue dont le siège est dans les mitochondries. Bien que plus lente que les fermentations, elle aboutit à une oxydation complète du substrat organique en CO2 et H2O : on dit que ce substrat est minéralisé (en effetle CO2 n'est pas une molécule organique). Le grand nombre de réactions mises en jeu explique une plus grande production d'ATP que les fermentations. Enfin, il est à noter qu'elle nécessite un apport de dioxygène O2.
3.2 La glycolyse, première étape des fermentations et de la respiration
La glycolyse consiste en une série de réactions anaérobies se déroulant dans le cytoplasme. Elle
consiste en l'oxydation du glucose qui donne finalement deux molécules d'acide pyruvique, ainsi que
deux molécules d'ATP et deux molécules de transporteurs réduits.
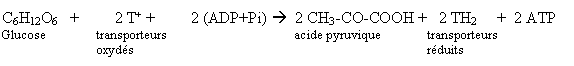
3.3 Mécanisme de la fermentation
Au cours de la fermentation, anaérobie, l'acide pyruvique est dégradé en une autre molécule organique, telle que l'éthanol (fermentation alcoolique) ou l'acide lactique (fermentation lactique). Deux nouveaux transporteurs réduits sont produits, mais il n'y a pas production supplémentaire d'ATP. Ces réactions sont donc faiblement énergétiques comparées à la respiration.
3.4 Mécanisme de la respiration
1) Le cycle de Krebs
Les réactions de respiration faisant suite à la glycolyse se déroulent dans les mitochondries, petits organites à double membrane dispersés dans le cytoplasme.
L'acide pyruvique pénètre dans la mitochondrie, où il subit une série de décarboxylations (= réactions conduisant à la libération de CO2) et de déshydrogénations. Les six molécules de carbone initialement présentes dans le glucose se retrouvent finalement dans des molécules de CO2 : il y a minéralisation du carbone organique.
Le mécanisme est le suivant :
- L'acide pyruvique subit une première décarboxylation / déshydrogénation
- Le groupement à deux carbone restant se fixe sur un composé en C4 (= à 4 atomes de carbone)
- Ce corps en C6 formé subit une série de réactions ( 2 décarboxylations et 4 déshydrogénations) à la fin desquelles est régénéré le corps en C4 initial : c'est le cycle de Krebs.
- Ensuite les transporteurs réduits produits pendant le cycle de Krebs mais aussi pendant la glycolyse sont réoxydés au niveau de la membrane interne de la mitochondrie : c'est la chaîne respiratoire.
2) La chaîne respiratoire.
La membrane interne des mitochondries comporte de nombreux replis ou crêtes mitochondriales, qui sont le siège de la chaîne respiratoire.
Celle-ci provient de la réoxydation des transporteurs réduits : les électrons libérés sont transportés par divers composés inclus dans la membrane interne de la mitochondrie. Pendant leur transport, ils favorisent le passage de protons (= ions H+) depuis la matrice (intérieur de la mitochondrie) vers l'espace intermembranaire. Il en résulte un gradient de protons (= une différence de concentration).
Le retour des protons vers la matrice peut libérer de l'énergie (car un gradient de protons implique l'existence d'une énergie potentielle) : le flux de protons est canalisé par une molécule particulière qui exploite cette énergie potentielle pour la synthèse d'ATP.
Ains les électrons parcourent la chaîne respiratoire jusqu'à un accepteur final : le dioxygène.
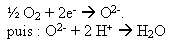
Bilan de la respiration :
Au cours de la respiration, la dégradation d'une molécule de glucose en CO2 conduit à la formation de 38 ATP :
2 proviennent de la glycolyse (voir plus haut) et 36 proviennent de la réoxydation des transporteurs dans la
mitochondrie.
4 Mots-clés et schéma récapitulatif
- métabolisme, réactions exoénergétiques, endoénergétiques ;
- ATP, ADP ;
- Fermentations, respiration ;
- Glycolyse, cycle de Krebs, chaîne respiratoire.