Génotype et phénotype
1 Introduction
L'information génétique dans la cellule est codée par l'ADN. Toutes les cellules d'un organisme donné possèdent la même quantité d'ADN, enroulé et compacté sous forme de chromosomes, groupés par paires chez les organismes diploïdes (animaux, végétaux…), et dont le nombre est particulier à une espèce donnée (23 paires chez l'homme).
La molécule d'ADN est constituée par un enchaînement de nucléotides ; ils sont constitués par un sucre, un groupement phosphate et quatre bases azotées différentes (A, T, G, C). L'enchaînement particulier de ces bases constitue l'information génétique, qui peut être transcrite puis traduite pour donner une protéine (voir fiches correspondantes). La portion d'ADN codant pour une protéine donnée est un gène. Le locus est l'endroit où se situe le gène sur le chromosome.
2 La notion d'allèles
2.1 Les allèles et les mutations
Les informations génétiques codées au niveau d'un même gène peuvent être différentes. Les différentes formes possibles d'un gène donné sont appelées les allèles. Cette diversité est rendue possible par la structure même des allèles. Ces derniers ne sont en effet que des séquences de nucléotides susceptibles d'être modifiées. Au niveau du seul nucléotide, on peut en effet déjà énoncer trois types de mutations possibles:
- On remplace un nucléotide de la chaîne par un autre: substitution
- On enlève un nucléotide de la chaîne: délétion
- On ajoute un nucléotide à la chaîne: insertion
Bien sûr, ces opérations de base peuvent se combiner à divers niveaux de la séquence nucléotidique pour aboutir à des modifications plus complexes. Chaque gène peut donc avoir plusieurs allèles différents et être ainsi polymorphe. On parle de polyallélisme.
2.2 Homozygotie, hétérozygotie
Le nombre d'allèles d'un même gène présent dans une cellule dépend du type de celle-ci :
- Si la cellule est une bactérie, elle ne possède qu'un seul chromosome ou en d'autres termes, qu'un seul allèle d'un gène donné.
- Si la cellule est haploïde (gamètes, nombreux champignons…), elle possède n chromosomes : il n'y a qu'un allèle par gène.
- Si la cellule est diploïde (elle possède 2n chromosomes réunis en paires dites d'homologues), chaque gène est présent en deux exemplaires, chacun sur un des deux chromosomes homologues. On en déduit qu'il y a donc deux allèles. Deux possibilités existent alors. Soit ceux-ci sont identiques et on dit de l'individu qu'il est homozygote pour le gène, soit ils sont différents et on parle de caractère hétérozygote pour le gène.
3 Expression des allèles et phénotypes
3.1 Phénotype et génotype
L'ensemble des allèles détenus par un individu constitue le génotype. Celui-ci reste avant tout une donnée génétique. En vis-à-vis de celui-ci, on place généralement le phénotype (ensemble des caractères observables d'un individu, par exemple la couleur des yeux).
Nous savons tout d'abord qu'un gène peut s'exprimer par la synthèse d'une protéine, et ce grâce au mécanisme de traduction de l'information génétique au niveau des ribosomes. Les protéines formées sont donc des produits directs de l'expression des allèles. A partir de deux allèles différents, le processus de synthèse protéique peut aboutir à :
- Des protéines fonctionnellement identiques (mutation muette ou neutre). C'est le cas lorsque la différence d'information portée par la séquence de nucléotides n'affecte pas la formule de la protéine (cela est rendu possible par le caractère redondant du code génétique). C'est aussi le cas quand la nouvelle séquence synthétisée n'a pas d'impact sur la fonction de la protéine.
- Des protéines différentes (mutation faux-sens ou non-sens). Certaines peuvent très bien, de par leur formule, ne pas être fonctionnelles : c'est très souvent le cas lorsqu'il y a insertion ou délétion d'un ou deux nucléotides, ou que la mutation implique la présence d'un autre acide aminé que celui initialement présent. Il peut aussi très bien ne pas y avoir de protéine du tout (rencontre d'un codon stop dès le début de la traduction).
Schéma récapitulatif
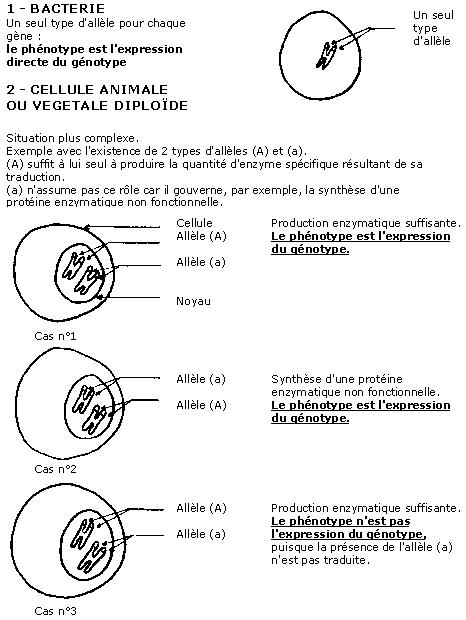
3.2 Dominance et récessivité
Lorsqu'un allèle donné s'exprime chez les individus homozygotes pour ce gène mais aussi chez les individus hétérozygotes, on dit qu'il est dominant.
Lorsqu'un allèle donné ne s'exprime que chez les individus homozygotes pour ce gène, on dit qu'il est récessif ; il ne s'exprime pas lorsque l'autre allèle du gène est dominant.
Enfin, lorsque deux allèles s'expriment tous les deux chez un individu hétérozygote pour le gène en question, on dit qu'ils sont codominants.
En tenant compte de ces remarques et si on considère chaque protéine comme expression à l'échelle moléculaire d'un phénotype, on explique aisément toute la diversité phénotypique par l'existence du polyallélisme (c'est à dire existence, dans une population, de nombreux allèles pour un seul gène).
4 La notion d'identité génétique
Un premier critère de distinction biologique est évidemment la notion d'espèce. Au sein d'une même espèce, la plus grande partie de l'information génétique est partagée: elle définit ce qu'on nomme les caractères de l'espèce. Cependant, à l'intérieur d'une espèce, au niveau des individus - et nous l'observons dans la diversité humaine- il existe des caractères propres (ex. couleur des yeux). Ceux-ci sont l'expression phénotypique de génotypes spécifiques.
Un exemple du programme permet d'illustrer cette diversité d'un point de vue biologique. A la surface de la membrane de ses cellules, chaque être humain possède en effet des combinaisons originales de glycoprotéines appelées marqueurs majeurs d'histocompatibilité ou plus communément marqueurs du Soi. Ceux-ci sont absolument différents entre individus et sont donc une véritable carte d'identité cellulaire. Là encore, la diversité des marqueurs du Soi est l'expression du polyallélisme de certains gènes. L'ensemble de ceux-ci constitue le complexe majeur d'histocompatibilité.
Mots-clés :
- génotype ; phénotype ;
- gène ; locus ; mutations : substitution, addition, délétion
- allèle : dominant, récessif, codominant ;
- diploïde ; homozygote, hétérozygote