Diversité génétique des populations et prévision en génétique humaine
1 La diversité génétique des populations
1.1 La définition d'une population et la fréquence allélique
Une espèce est définie comme un ensemble d'individus pouvant se reproduire entre eux et dont les descendants sont fertiles. Au sein d'une espèce, on désigne par le terme de population, un ensemble d'individus dans un milieu écologique donné et se reproduisant entre eux. On peut définir pour une population, à un instant donné, la pléiade d'allèles qu'elle possède et la fréquence relative de chacun.
Une population est panmictique lorsque l'union des individus reproducteurs se fait de façon aléatoire. Cela implique un nombre infini de reproducteurs. Si la panmixie est respectée, les fréquences génotypiques et alléliques restent stables. Il existe de nombreux cas cependant pour lesquels les fréquences alléliques varient au cours du temps : dans le cas où l'union des reproducteurs ne se fait plus au hasard (par exemple, petite population) ou dans le cas où un génotype devient supérieur aux autres (un génotype allonge la longévité des individus).
1.2 La sélection naturelle
L'incidence de la sélection naturelle et des mutations est négligeable à l'échelle de quelques générations, mais est fondamentale à l'échelle des temps géologiques.
- La sélection naturelle a un impact important sur l'évolution de la diversité au sein de la population. En particulier, on peut souligner deux cas de figure : la sélection naturelle est diversifiante lorsqu'elle favorise le génotype hétérozygote; elle est uniformisante si elle favorise le génotype homozygote. Les facteurs sélectifs sont variés. Il peut s'agir de facteurs physico-chimiques naturels (comme la température, l'hygrométrie, le pH, la salinité) ou des facteurs biotiques : ainsi les prédateurs pratiquent une sélection en tuant les proies les moins protégées. La concurrence inter et intra-spécifique pour l'occupation de l'espace vital ainsi que la recherche de la nourriture et du partenaire sexuel interviennent aussi dans la sélection naturelle.
- Les mutations permettent alors de compenser l'élimination de certains génotypes due à la sélection uniformisante. Dans le cas de la sélection diversifiante, elles décalent les équilibres de fréquence allèlique.
L'action combinée de la sélection naturelle et des mutations tend à maintenir les fréquences alléliques de la population à une valeur d'équilibre pour des populations de tailles conséquentes occupant un milieu stable.
1.3 La notion de dérive génétique et de divergence génétique
Dans une population de taille réduite, on assiste à une variation aléatoire des fréquences alléliques d'une génération à une autre. La dérive génétique est le phénomène de réduction du polymorphisme génétique au sein d'une petite population : l'écart génétique entre les individus de la population diminue. Si au contraire, à partir d'une population mère, deux populations filles s'isolent, la dérive génétique s'effectuera de façon indépendante dans chacune d'elles. La différence entre les pools génétiques augmente entre ces populations, c'est la divergence génétique. Les deux sous populations peuvent diverger jusqu'au point où les individus appartenant à chacune d'elles ne sont plus inter-féconds. Il apparaît ainsi une nouvelle espèce, c'est le mécanisme de spéciation. Les facteurs conduisant à la séparation d'une population en deux sous-populations sont nombreux : ils peuvent être physiques (barrière naturelle), comportementaux (individus plus actifs la nuit et d'autres plus actifs le jour), biologiques (préférences sexuels qui séparent les reproducteurs).
2 Les prévisions en génétique humaine
2.1 Position du problème
La génétique humaine cherche à évaluer la probabilité de phénotypes déterminés. Les contraintes sont cependant nombreuses lorsque l'on étudie l'espèce humaine : la fécondité est faible, le temps séparant deux générations est élevé et l'expérimentation est, bien entendu, impossible. Pour remédier à ces contraintes, l'une des solutions adoptées est l'étude de l'arbre généalogique de l'individu dont on recherche le génotype. On recense alors le phénotype de tous les membres d'une famille donnée. Cette méthode est souvent utilisée lorsque l'on souhaite tracer l'allèle responsable d'une maladie qualifiée alors de maladie héréditaire.
Dans la plupart des problèmes présentés, on s'intéressera donc à un allèle responsable de l'apparition d'une maladie. On cherchera à déterminer la probabilité pour les descendants d'une famille d'être atteints en étudiant l'arbre généalogique de la famille. Pour répondre à cette question, il faut déterminer deux points essentiels :
- L'allèle responsable de la maladie est-il récessif ou dominant ?
- L'allèle responsable de la maladie est-il porté par les autosomes (22 paires de chromosomes homologues chez l'espèce humaine) ou les chromosomes sexuels (X et Y) ?
2.2 Allèle autosomal dominant ou récessif ?
Dans le cas d'une maladie ou une anomalie liée à un allèle dominant porté par un autosome, la particularité s'exprime chez tous les porteurs de l'allèle morbide, même hétérozygotes. Les probabilités pour que la descendance soit affectée sont de 100% si le parent porteur est homozygote, de 75% si les deux parents sont hétérozygotes, et de 50% avec un seul parent porteur hétérozygote.
Dans le cas d'une maladie ou d'une anomalie liée à un allèle récessif porté par un autosome, la particularité n'apparaît que si l'individu est homozygote.
- Deux parents homozygotes transmettent toujours la maladie ou l'anomalie considérée.
- Deux parents hétérozygotes, ne présentant pas les symptômes de la maladie ou de l'anomalie,
ont une probabilité de :
- ½ pour que les gamètes soient porteurs
- ¼ pour avoir un enfant homozygote
2.3 Tableaux de croisement dans le cas d'un allèle autosomal dominant
Tableaux de croisement dans le cas d'un allèle autosomal dominant
M : allèle dominant malade; n : allèle récessif normal
| Père et mère hétérozygotes | ||
|---|---|---|
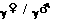 |
n | M |
| n | n/n | M/n |
| M | M/n | M/M |
| Probabilité d'avoir un enfant atteint : ¾ | ||
| L'un des parents est homozygote, l'autre hétérozygote | ||
|---|---|---|
|
n | M |
| M | M/n | M/M |
| n | n/n | M/n |
| Tous les enfants sont atteints | ||
2.4 Tableaux de croisement dans le cas d'un allèle autosomal récessif
Tableaux de croisement dans le cas d'un allèle autosomal récessif
N : allèle dominant normal; M : allèle récessif malade
| Père et mère hétérozygotes | ||
|---|---|---|
|
N | m |
| N | N/N | N/m |
| m | N/m | m/m |
| Probabilité d'avoir un enfant atteint : ¼ | ||
| L'un des parents est homozygote, l'autre hétérozygote | ||
|---|---|---|
|
m | N |
| N | m/N | N/N |
| N | m/N | N/N |
| Aucun enfant n'est atteint | ||
2.5 Allèle porté par un chromosome sexuel
- Chromosome Y : Dans le cas d'une maladie liée à un allèle récessif porté par le chromosome sexuel Y, la maladie apparaît chez les garçons porteurs de l'allèle. Les pères porteurs de l'allèle le transmettent à leur fils.
- Chromosome X : Dans le cas d'une maladie liée à un allèle récessif porté par le chromosome sexuel X, la maladie apparaît, d'une part, chez les filles homozygotes et, d'autre part, chez les garçons porteurs de l'allèle sur leur chromosome X.
- Si une mère est homozygote, tous ses fils présentent la maladie.
- Si une mère est hétérozygote, la probabilité pour que ses fils soient atteints est de ½.
- Dans les deux cas, la probabilité pour que les filles portent l'allèle récessif est de ½.